Brain connectivity
As an undergraduate physics student, I wondered why those bulky neuroscience textbooks of my friends at medicine school devoted so many pages to describe the gray matter – a layer of tissue one millimetre thin – but only dedicated only a handful pages to the white matter, a tissue that occupies about half of the volume of the brain.
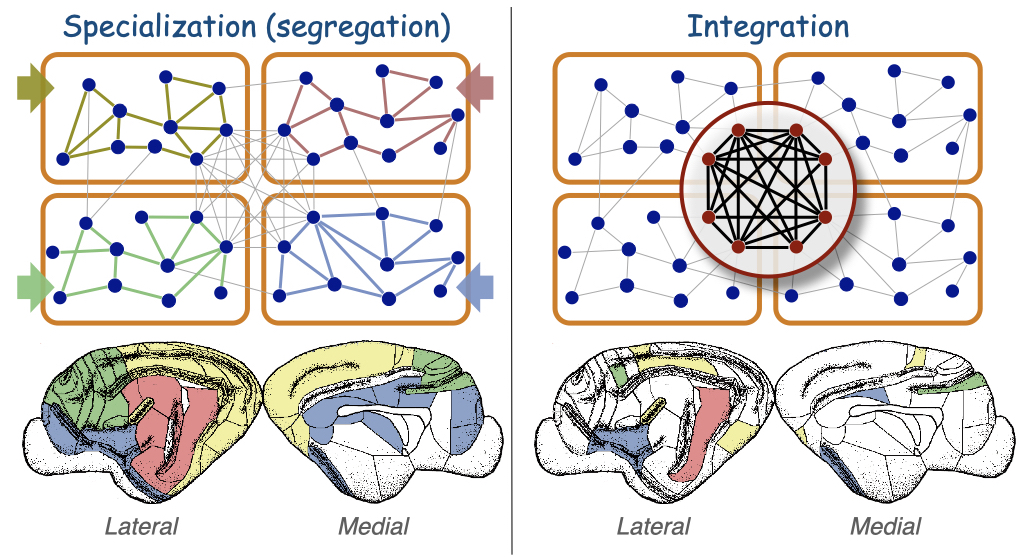
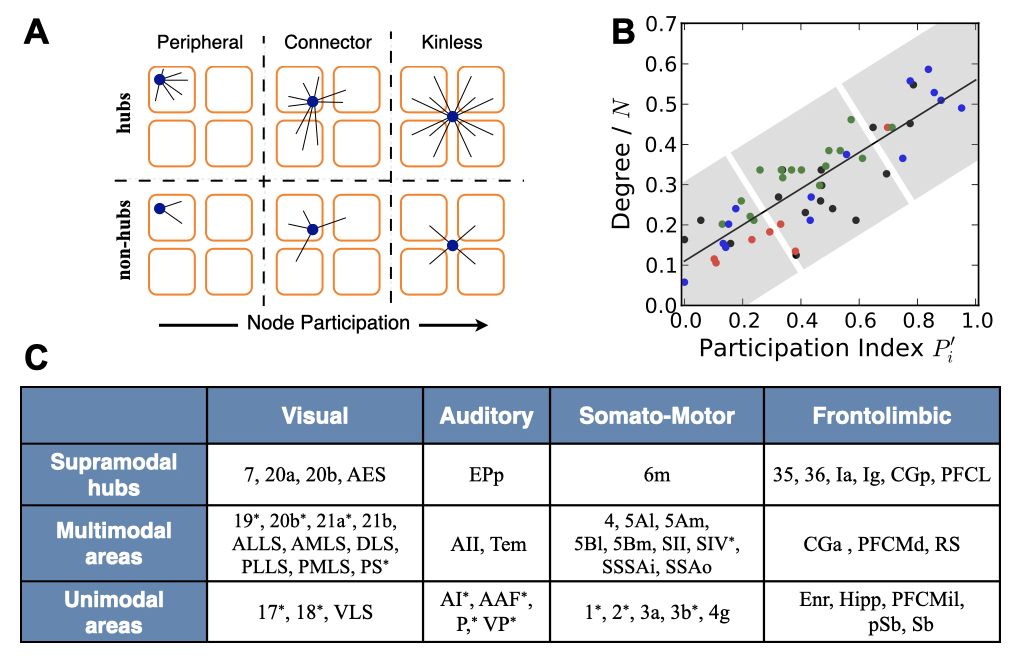
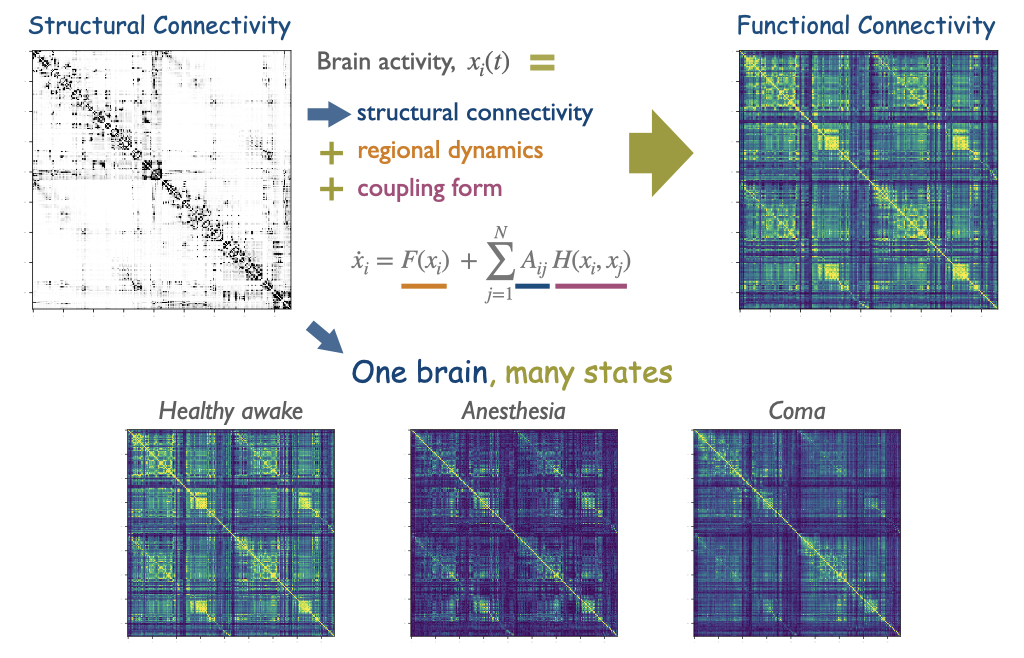
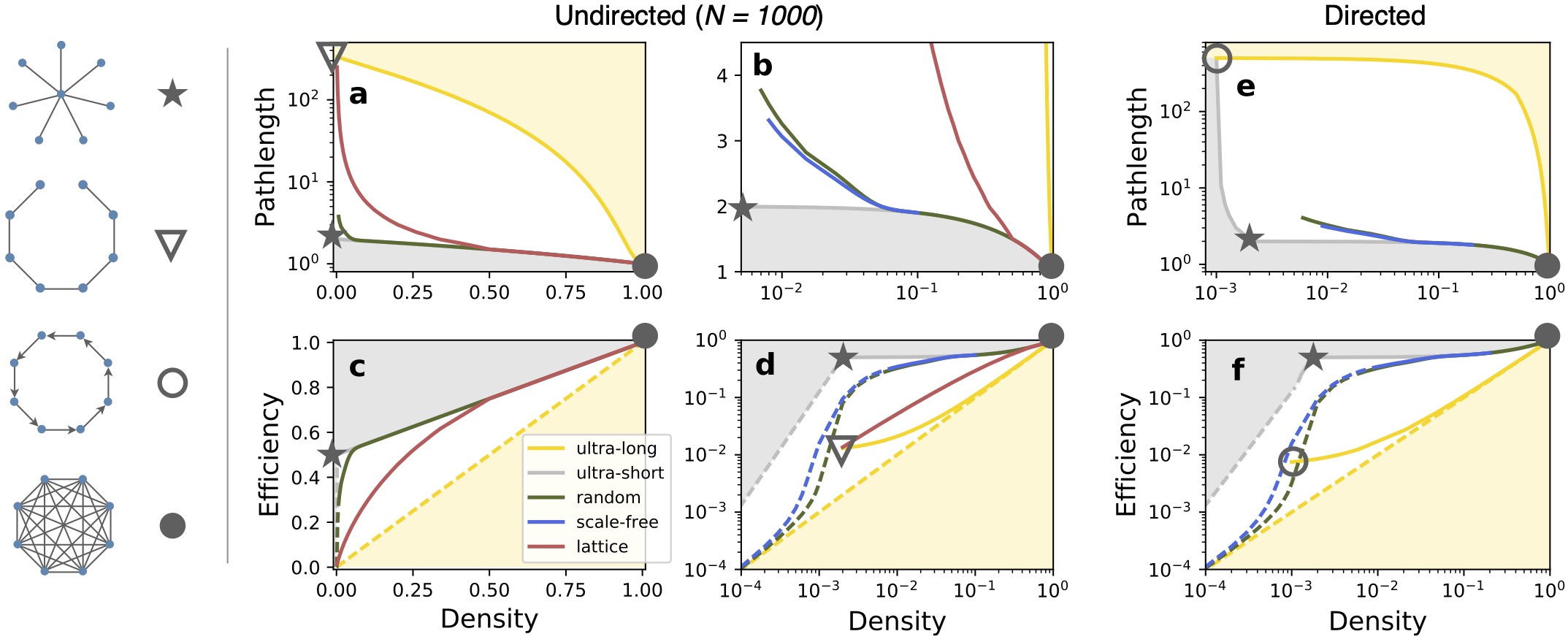
Today, we know that the white matter is the “internet of the brain“: a vast network of cables that allows the communication between different parts of the brain and cortex. My work has helped to better understand this intricate network of communications. Building upon previous seminal works, and based on the cortico- cortical connectivity of cats, we identified that cortico-cortical networks have a modular but centralised architecture, with few regions – the rich-club – centralising those communication paths.